혈관내피성장인자(vascular endothelial growth factor, VEGF)는 신생혈관생성의 중요한 매개체로서 주로 혈관내피세포에서 발현되는 특이 수용체에 결합하여 내피세포의 증식과 이동, 세포외기질의 분해와 같은 신생혈관생성작용을 나타낸다[1]. VEGF는 당뇨망막병증과 망막정맥폐쇄에 의한 신생혈관을 촉진하는 작용을 나타내는데, 임상적으로 신생혈관 억제 등을 목적으로 항혈관내피성장인자(anti-VEGF)로 만든 약제를 사용하고 있으나 부작용의 하나로 안압상승도 나타날 수 있다[2-4]. VEGF는 혈액뇌장벽의 투과도와 혈관내피세포의 투과도를 증가시키는 작용을 나타내며[5,6], 혈관내피세포와 평활근세포에서 VEGF의 작용기전으로 nitric oxide (NO)가 관여하는 것으로 알려져 있다[7,8]. 또한 안구 내에 조직과 망막혈관내피세포에서도 VEGF는 투과도를 증가시키며 이 과정에서도 NO가 관여한다고 알려져 있다[9,10].
섬유주는 방수유출로의 조절에 중요한 역할을 하는데, 섬유주를 통한 방수유출로의 저항이 증가되어 개방각녹내장을 유발하는 것으로 알려져 있다[11,12]. 섬유주를 통한 방수 유출에서 섬유주세포는 세포외기질의 대사를 조절하여 섬유주의 저항을 조절함으로써 안압을 유지하는 중요한 역할을 한다[13,14]. VEGF는 세포외기질을 분해하고 조절하는 역할을 하는 기질금속단백분해효소(matrix metalloproteinase, MMP)의 활성을 증가시키는데, 혈관내피세포에서 MMP-1과 MMP-2의 분비를 증가시키며 평활근세포에서도 MMP의 발현을 증가시킨다[15-17]. 섬유주세포는 혈관내피세포와 평활근세포의 성질을 나타내므로[18] anti-VEGF는 섬유주에서 MMP의 분비를 저하시켜 세포외기질의 축척을 유발하여 섬유주를 통한 방수유출을 저하시킬 가능성이 있을 것이나, 섬유주에서 anti-VEGF가 섬유주의 유출에 미치는 영향과 섬유주를 통한 방수유출을 촉진하는 작용을 나타내는 MMP에 대한 영향은 아직 자세히 밝혀지지 않았다. 이에 따라 본 연구에서는 배양한 사람의 섬유주세포를 이용하여 anti-VEGF가 섬유주의 투과도에 미치는 영향을 알아보고 세포외기질의 대사에 중요한 역할을 하는 MMP-1/-2의 분비에 미치는 영향을 알아보고자 하였다.
대상과 방법
세포배양 및 약물처리
본 연구는 대구가톨릭대학병원 의학윤리심의위원회(Institutional Review Board, IRB)의 승인을 받았고(IRB 승인 번호: CR-19-059-L) 헬싱키선언을 따라 시행되었다. 기존의 안병력이 없는 안구이식에 동의한 50세 환자로부터 안구은행에서 얻은 사후 6시간 이내의 안구에서 전방각에서 섬유주를 벗겨내어 배양접시에 옮긴 후 항생제(antibiotic antimycotic solution containing penicillin, streptomycin and amphotericin B, Gibco, Invitrogen, Carlsbad, CA, USA)와 10% 우태아혈청(fetal bovine serum, FBS) (HyClone, Cytiva, Global Life Sciences Solutions USA LLC, Marlborough, MA, USA)이 포함된 Dulbecco’s modified Eagle’s medium (DMEM; Gibco, Invitrogen, Carlsbad, CA, USA) 배지를 사용하여 37°C, 5% CO2 배양기에서 초대배양하였다. 섬유주세포가 이식된 조직편 주위로 자라 나오는 양상과 형태학적으로 섬유주세포의 특징을 확인한 후 섬유주 조직의 이식편을 제거하고 배양을 계속하였으며 세포가 배양접시에 충만해지면 1:3의 비율로 트립신 처리하여 계대배양하였다. 배양된 세포를 섬유주세포 표지자(fibronectin, lamininm, vimentin)로 확인한 후 동결보존하였다. 실험을 위하여 동결보존한 세포를 해동한 후 배양한 다음 인체의 섬유주세포에 anti-VEGF 제제인 bevacizumab (BV; Avastin®, Genentech, South San Francisco, CA, USA)을 0.25, 0.50 mg/mL의 농도로 24시간 동안 노출시켰다. 이때 BV의 농도는 0.1% FBS가 포함된 DMEM 배지를 이용하여 희석하여 조절하였으며 대조군은 동량의 0.1% FBS가 함유된 DMEM을 사용하였다.
MTT assay
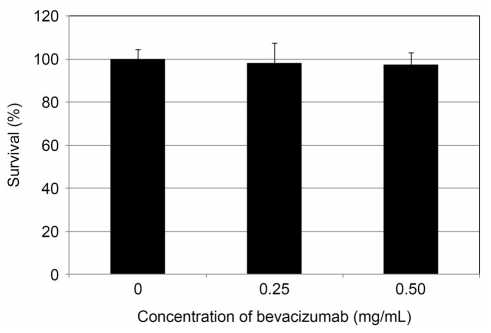
세포의 생존에 대한 효과는 3-[4, 5 -dimethylthiazol-2-yl]-2, 5-diphenyltetrazolium bromide (MTT; Sigma-Aldrich, MO, USA) assay를 이용하였으며 세포의 생존 정도는 실험군의 값을 약물처리를 하지 않은 대조군의 비로 나누어 백분율로 나타내었다.
Carboxyfluorescein permeability assay
Transwell (Pore size 0.3 ❍m, polycarbonate membrane, #3460, Corning Life Sciences, Tewksbury, MA, USA)의 내측 chamber에 세포가 단일 세포층으로 충만하게 자란 것을 확인한 후 BV에 24시간 노출시킨 후 투과도 검사를 시행하였다[19-21]. 내측 chamber에 자라고 있는 세포를 세척한 다음 50 mm carboxyfluorescein (Sigma, St. Louis, MO, USA)을 노출시키고 노출 2시간 후 외측 chamber로 투과된 carboxyfluorescein의 농도를 532 nm에서 분광형광계(Fluostar Optima, BMG Labtech, Ortenberg, Germany)로 측정하여 백분율로 나타내었다.
Trans-endothelial electrical resistance (TEER) assay
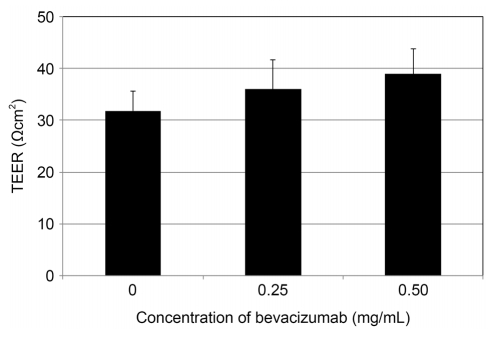
섬유주를 통한 투과도를 측정하기 위한 또 다른 방법으로 TEER assay를 시행하여 섬유주단층세포층의 저항도를 측정하였는데[22,23] TEER assay는 단층 세포층에서 전기적 저항이 증가할수록 투과도가 감소하는 원리를 이용한 방법이다. 섬유주세포를 내측 chamber에 충만하게 배양한 다음 BV에 24시간 노출시킨 후 TEER을 epithelial voltohmmeter (EVOM [2]; World Precision Instruments, Sarasota, FL, USA)를 이용하여 측정하였으며 그 결과를 net value (Ωcm2)로 기록하였다.
MMP-1/-2 단백의 활성 측정을 위한 Western blot
단백질 추출에는 RIPA buffer (Thermoscientific, Carlsbad, CA, USA)를 사용하였다. Cell lifter (Corning, Acton, MA, USA)로 세포를 모아 원심 분리하여 상층액을 모아 -80°C에 보관하였다. 단백질량은 BCA protein assay reagent kit (Thermoscientific, Carlsbad, CA, USA)를 사용하여 측정하였다. 정량한 샘플을 같은 양으로 NuPAGE 4-12% Bis-Tris gel (Invitrogen, Carlsbad, CA, USA)에 전기영동한 후 Xcell Surelock (Invitrogen, Carlsbad, CA, USA)을 이용하여 니트로셀룰로오스 막(Invitrogen, Carlsbad, CA, USA)에 1시간 동안 옮긴 다음 막 차단 용액(Invitrogen, Carlsbad, CA, USA)에서 1시간 동안 정치배양하였다. Goat anti-rabbit horseradish peroxidase-conjugated된 이차 항체(Santacruz Biotechnology Inc., Dallas, TX, USA)를 막 차단 용액에 1:3,000으로 희석하여 처리하고 상온에서 1시간 반응시켰다. 항체를 반응시킨 막을 SuperSignal West Pico Chemiluminescent Substrate (Thermoscientific, Carlsbad, CA, USA)를 이용하여 Gel Doc XR+ (Bio-rad, Hercules, CA, USA)로 단백의 발현양을 확인하였다. 이때 glyceraldehyde 3-phosphate dehydrogenase를 internal standard로 사용하였다.
Griess assay
BV이 섬유주세포에서 내인성 일산화질소의 생성에 미치는 영향을 알아보기 위하여 Griess assay를[24] 시행하여 배지에서의 nitrite 생성량을 측정하였다. 약물처리한 세포 배지에 동량의 Griess 반응액(Sigma, St. Louis, MO, USA)을 섞은 후 96-well plate에 옮겨 NO 생성의 반응물인 아질산염(nitrite)의 양을 spectrophotometer로 540 nm에서 흡광도를 측정하여 정량하였다. 이때 표준치를 구하기 위해 sodium nitrite (Sigma, St. Louis, MO, USA)를 단계적으로 희석하여 사용하였다.
결 과
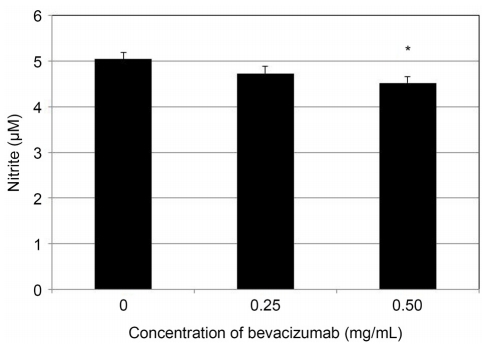
BV가 NO의 생성에 미치는 영향
약제에 노출되지 않은 대조군에 비하여 0.25 mg/mL BV는 배지에서 nitrite의 생성에 유의한 영향을 미치지 않았으나(p=0.190), 0.50 mg/mL BV는 배지에서 nitrite의 생성량을 유의하게 증가시켰다(p=0.023) (Fig. 6).
고 찰
Anti-VEGF는 재조합 단일클론 IgG 항체이며 안구 조직에 미치는 영향은 이미 잘 알려져 있다[17]. 임상적으로 사용되는 농도에서는 anti-VEGF는 독성을 유발하지 않는다는 보고도 있으나 고농도에 노출될 경우 섬유주세포에 대한 독성을 유발할 수 있다[25-28]. 그러나 본 실험에 사용된 저농도의 anti-VEGF는 기존 연구 결과와 마찬가지로 섬유주세포 생존에 유의한 영향을 미치지 않았다[25].
Anti-VEGF 제제의 부작용의 하나로 안압상승이 초래될 수 있는데[2-4], 안압상승의 기전은 아직 명확하게 밝혀지지 않았으나 그 기전의 하나로서 anti-VEGF는 NO의 생성을 억제하는 작용이 있으므로 섬유주에 노출될 경우 방수유출을 억제함으로써 안압상승을 유발하는 기전으로 작용할 가능성이 제시된 바 있다[27].
Anti-VEGF가 방수유출에 미치는 영향을 실험실 내 연구로 알아보기 위하여 시행한 anti-VEGF의 일종인 BV을 사용한 섬유주단층세포층의 투과도검사에서 0.50 mg/mL BV는 carboxyfluorescein의 투과도를 유의하게 감소시켰다. 따라서 anti-VEGF는 농도가 높을수록 섬유주세포의 투과도를 감소시킬 가능성이 있다.
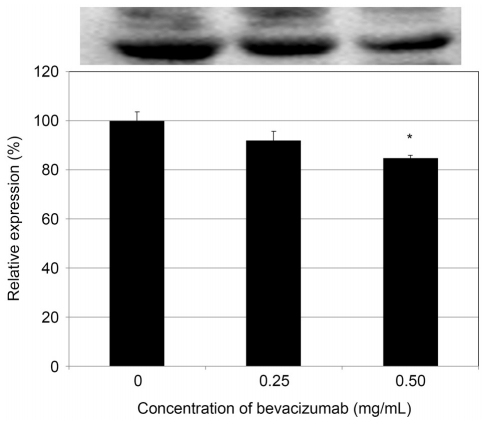
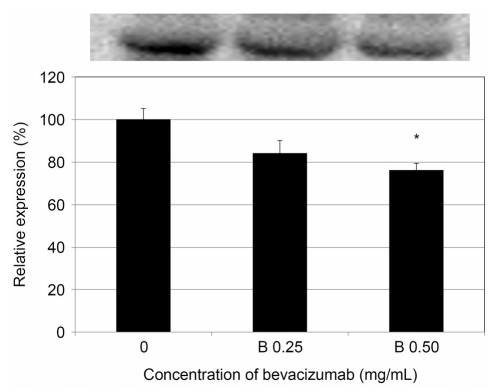
섬유주를 통한 방수유출의 저항을 조절하는 데에 있어서 MMP에 의한 세포외기질의 대사조절이 중요한 역할을 한다. 섬유주세포와 조직에서 다양한 종류의 MMP가 발현되는데 이들 MMP 중 MMP-1, 3, 9은 정상 상태에서는 발현이 미약하나 다양한 자극에 의해 발현이 증가되며, MMP-2는 비교적 많이 발현되는 것으로 알려져 있으므로[13,14] 본 연구에서는 MMP-1과 MMP-2의 발현을 조사하였다. VEGF는 혈관내피세포에서 MMP-1과 MMP-2의 분비를 증가시키며, 평활근세포에서도 MMP의 발현을 증가시키므로[15-17], anti-VEGF는 MMP의 활성을 감소시켜 세포외기질의 대사를 저해함으로써 섬유주의 저항을 증가시킬 가능성이 있으나, 인체의 섬유주세포에서 anti-VEGF가 섬유주세포의 MMP 활성에 미치는 영향은 아직 알려지지 않았다. 이에 따라 시행한 본 연구의 결과에서 BV는 MMP-1/-2의 활성을 각각 억제하였다. 따라서 anti-VEGF의 농도가 증가할수록 MMP 단백의 활성을 저하시켜 세포외기질의 축척을 유발하여 방수유출에 대한 섬유주의 저항을 증가시킬 수 있을 것이다[29,30].
VEGF가 투과도를 증가시키는 기전의 하나로 NO가 관여하는 것으로 알려져 있다[5-9,31]. NO는 섬유주를 이완시켜 방수유출을 증가시키는 작용을 나타내므로[32-34] anti-VEGF가 NO의 생성을 저하시킨다면 방수유출을 저하시킬 수 있을 것이다. MMP는 조직과 병적 상태에 따라 매우 다양한 작용을 나타내는데[35] NO와 MMP의 발현에 관한 상호작용에 대해서는 세포의 종류에 따라 다양한 결과가 보고되어 있으며 NO는 MMP-1의 발현을 촉진하기도 하고 MMP-2의 발현을 억제하기도 한다[36-38]. 이를 규명하기 위하여 BV을 이용하여 NO의 생성에 미치는 영향을 조사한 결과, 섬유주에서는 anti-VEGF가 NO의 생성을 억제하는 것으로 나타났으며[27] 이와 동반하여 MMP의 발현이 저하되었다. 이러한 결과로 보아 섬유주세포에서는 NO가 MMP의 발현을 저해할 가능성도 있을 것으로 생각된다. 본 연구에서는 MMP-1/-2의 발현에 미치는 영향을 조사하였는데 섬유주에서는 다양한 MMP와 그 저해제가 발현되므로[30,39] 여러 종류의 MMP에 미치는 영향과 NO와 MMP의 상호작용에 대해서는 향후 보다 자세한 연구를 요한다.
조직에서 용액의 투과경로는 세포경유경로와 세포주변 경로를 통해 투과되는데[40], 본 연구에서 BV는 세포주변경로를 통한 투과성을 나타내는 섬유주단층세포층의 carboxyfluorrescein 투과도를 감소시켰으나 섬유주단층세포층의 저항도에는 유의한 영향을 미치지 않았다. 그러나 섬유주에서는 세포주변경로를 통한 방수의 유출은 미미하며 주로 세포경유경로인 거대수포를 통해서 방수가 유출되므로[41-43] BV가 세포주변경로를 통한 유출을 시킨다는 본 연구의 실험실 내 결과는 실제 생체 내 섬유주 조직과의 조건은 다를 수 있다. 또한 두 경로는 서로 밀접하게 연관되어 조절될 뿐만 아니라[10,44] anti-VEGF가 세포경유경로를 통한 투과도를 증가시킨다는 보고도 있으므로[45], anti-VEGF가 세포경유경로에 미치는 영향을 완전히 배제할 수는 없을 것이며 이에 관해서는 향후 보다 자세한 연구를 요한다. 또한 본 실험에서는 임상적으로 사용 중인 BV를 희석하여 사용하였으므로 BV에 함유되어 있는 다양한 첨가물의 영향도 고려할 필요가 있을 것이다. 또한 anti-VEGF를 인체의 유리체내에 주사하는 경우 다양한 기전에 의해 안압이 상승할 수 있는 점도 고려해야 할 것이다[46]. 결론적으로 anti-VEGF는 섬유주세포의 투과도를 감소시켰으며, NO의 생성 저하와 함께 MMP의 분비를 감소시킴을 알 수 있었다.


 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Download Citation
Download Citation Print
Print